|
Росина В. В., 2004. Голоценовые рукокрылые
(Chiroptera, Mammalia) и особенности их тафономии в условиях пещер
Северо-Западного Алтая. - Экосистемные перестройки и эволюция биосферы,
вып. 6. М., ПИН РАН, С. 110-118.
ГОЛОЦЕНОВЫЕ РУКОКРЫЛЫЕ (CHIROPTERA, MAMMALIA) И
ОСОБЕННОСТИ ИХ ТАФОНОМИИ В УСЛОВИЯХ ПЕЩЕР СЕВЕРО-ЗАПАДНОГО АЛТАЯ
В.В. Росина
Палеонтологический Институт РАН
В голоценовых сборах из пещер северо-западного
Алтая обнаружены остатки девяти видов рукокрылых: Myotis daubentonii,
M. brandtii, M. dasycneme, М. frater, M.
ikonnikovii, M. blythii, Plecotus auritus, Eptesicus
nilssonii и Murina leucogaster. Самыми многочисленными
и обычными оказались кости М. daubentonii, M. brandtii
и М. dasycneme. Наиболее редки находки М. frater.
На процесс захорония остатков рукокрылых заметное влияние оказывают
видовая избительность мест локализации животных и особенности их
морфологии.
V.V. Rosina Holocene bats (Chiroptera, Mammalia) and their taphonomic
peculiarities in the caves of North-West Altai
In the Holocene sediments from Altai caves the following 9 bats
species discovered were: Myotis daubentonii, M. brandtii,
M. dasycneme, M. frater, M. ikonnikovii, M.
blythii, Plecotus auritus, Eptesicus nilssonii and
Murina leucogaster. The first tree species are the most common
and numerous. Myotis frater is the rarest species. The peculiarities
of the bats species localizations in the caves and their morphology
are very important for the taphonomical analysis.
ВВЕДЕНИЕ
Таксономическое разнообразие современных рукокрылых
в значительной мере определяется структурой их сообществ в предшествующие
эпохи. Важнейшим звеном в истории летучих мышей Алтая является их
голоценовое сообщество, которое до настоящего времени оставалось
крайне малоизученным.
Голоценовые кости рукокрылых зачастую в массовом количестве встречаются
в пещерных отложениях Алтая. Одни из первых упоминаний о голоценовых
остатках рукокрылых из алтайских пещер можно найти в работе П.П.
Стрелкова (1968). Позднее сборами субфоссильного материала по летучим
мышам в пещерах Алтая занимался Н.Д. Оводов (1972, 1974, 1999).
Традиционно считается, что накопление остатков рукокрылых в пещерах
происходит преимущественно в ходе естественной гибели зверьков из
состава зимующих популяций (Оводов, 1974; Тиунов, 1997; Филиппов,
Тиунов, 1999).
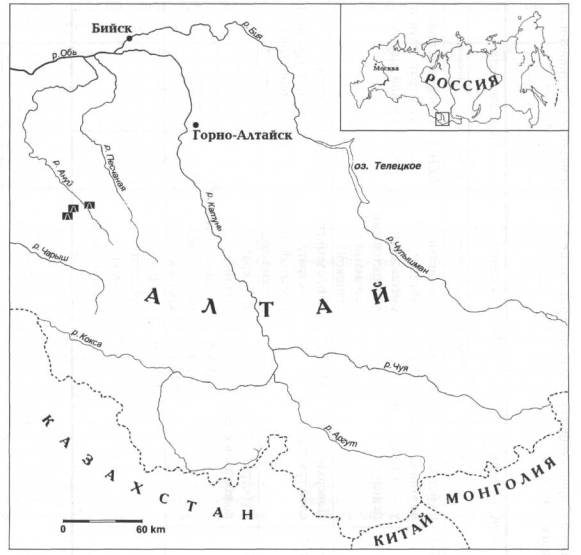
Рис.
1. Район исследований: бассейн р. Ануй северо-западного Алтая (рис.
А.В. Абдульмановой).
По способам концентрации костного материала все
пещерные захоронения можно разделить на три типа. Первый тип –
карстовые щели и трещины, накопление костного материала в осадках
которых происходило без участия биогенного фактора. Как правило,
это протяженные карстовые воронки, в донных известняках которых
посредством грунтовых вод происходит накопление костей. Второй тип
– скальные ниши и пещеры, в осадочном материале которых происходила
концентрация костей из погадок хищных птиц. Чаще всего эти местонахождения
имеют голоценовый и позднеплейстоценовый возраст. Третий тип –
пещерные стоянки древнего человека. В этом случае кости крупных
млекопитающих в пещеры приносил сам человек, а костные остатки мелких
животных поступали в пещерную полость из экскрементов собачьих,
сопровождавших человека (Агаджанян, 1987).
Накопление осадочного материала внутри пещер происходит медленно
и неравномерно. Это отражается в замедленности тафономических процессов
и в некоторых случаях может привести к полному разрушению костного
материала. Например, растворению его в лужах пещерной воды, имеющей
довольно агрессивный химический состав, т.к. холод стимулирует кислую
среду пещерных вод. Содержание СО2 в водах известковых пород почти
в 2700 раз выше, чем в дождевой. В подземных условиях углекислота
ведет себя более активно и во многом определяет химический состав
пещерной воды. Конденсационные воды, просачиваясь через отложения
пещеры, способны растворять карбонаты до бикарбонатов, а при повышении
температуры опять отлагать карбонаты (Николаев, 1994).
Наше исследование имело две основные цели: во-первых, изучить голоценовое
сообщество летучих мышей пещер северо-западного Алтая, и, сравнив
его с современным, выявить динамику произошедших изменений; во-вторых,
понять механизмы начальных этапов фоссилизации рукокрылых в пещерных
условиях.
МАТЕРИАЛ И МЕТОДЫ
На территории северо-западного Алтая в четырех
пещерах: Музейная, Титюниха, Разбойная и Денисова, расположенных
в бассейне р. Ануй (рис. 1), нами были собраны субфоссильные остатки
рукокрылых.
В поверхностном слое грунта (не глубже 10 см) в местах скопления
костей мелких животных, а также в трещинах стен пещер были взяты
пробы субстрата (табл. 1). Чаще всего субстрат представлял собой
смесь костного материала, глины или мелкой кальцитовой крошки.
| Пещ. Разбойная |
Пещ. Титюниха |
Пещ. Музейная |
Денисова пещ. |
| Обозн. |
Описание |
Обозн. |
Описание |
Обозн. |
Описание |
Обозн. |
Описание |
| РАЗ-1 |
Около 30 м от входа. Вынос водного потока вдоль тектонического шва |
ТИТ-1 |
Субстрат из ложбины в стене у входа в пещеру |
МУЗ-1 МУЗ-3 |
Глинистый субстрат, смешанный с кальцитовыми призмами и крошкой.
|
ДЕН-1 |
Поверхностный субстрат
грунта правой галереи |
| n |
n
|
ТИТ-2 ТИТ-4 |
Поверхностный слой грунта |
МУЗ-2 |
Черноземный субстрат,
намытый водой в узкий ход |
ДЕН-2 |
Субстрат из лужи в трещине в стене (правая галерея) |
| n |
n |
ТИТ-3 |
Субстрат из ложбины в стене |
МУЗ-4 МУЗ-5 МУЗ-6
МУЗ-7 МУЗ-8 |
Субстрат
стенных трещин: смесь кальцитовой
крошки |
n |
n |
| n |
n |
n |
n |
МУЗ-9 |
Осыпной субстрат: обломки кальцитовых наростов |
n |
n |
Таблица
1. Характеристика проб сборов субфоссильных материалов по летучим
мышам из голоценовых отложений пещер северо-западного Алтая.
|
пещеры
виды
|
Музейная
|
Разбойничья
|
Титюниха
|
Денисова
|
|
1
|
2
|
3
|
1
|
2
|
3
|
1
|
2
|
3
|
1
|
2
|
3
|
|
Myotis
daubentonii
|
17
|
54,8
|
75
|
5
|
41,7
|
11
|
16
|
48,5
|
24
|
2
|
66,7
|
6
|
|
M. brandtii
|
11
|
35,5
|
48
|
4
|
33,3
|
9
|
4
|
12,1
|
6
|
—
|
0,0
|
n
|
|
M. ikonnikovii
|
3
|
9,7
|
13
|
3
|
25,0
|
6
|
10
|
30,3
|
15
|
1
|
33,3
|
3
|
|
M.
frater
|
—
|
0,0
|
n
|
—
|
0,0
|
n
|
3
|
9,1
|
4
|
—
|
0,0
|
n
|
|
всего1
|
31
|
100
|
n
|
12
|
100
|
n
|
33
|
100
|
n
|
3
|
100
|
n
|
|
всего2
|
105
|
n
|
136
|
14
|
n
|
26
|
16
|
n
|
49
|
6
|
n
|
9
|
Таблица
2. Видовой состав рукокрылых в голоценовых отложениях пещер северо-западного
Алтая.
При отмывке были использованы сита с размером ячеи
0,5 мм. Рукокрылые представлены как краниальными, так и посткраниальными
остатками. Из первых присутствуют черепа различной степени поврежденности,
сильно поврежденные верхнечелюстные кости, целые и фрагментарные
части нижних челюстей с зубами и без, разрозненные верхние и нижние
зубы (резцы, клыки, премоляры, моляры), а также барабанные кости.
Посткраниальные остатки представлены трубчатыми костями конечностей,
лопатками, тазовыми костями, разрозненными костями осевого скелета.
Основная масса остатков без заметных следов фоссилизации, лишь некоторые
покрыты корочкой современного натечного кальцита белого, серого
и грязно-желтого цвета. Цвет остеологического материала преимущественного
белый, реже желтоватый, очень редко – рыжий. Видовая идентификация
проводилась только для краниальных остатков. Всего было просмотрено
около 1000 костей рукокрылых, из них определено 378 краниальных
остатков (табл. 2). При идентификации голоценовых сборов были использованы
сравнительные остеологические материалы по современным летучим мышам
территории северо-западного Алтая.
Для анализа структуры голоценового сообщества рукокрылых были рассчитаны:
встречаемость – процентная доля числа пещер, в которых были
обнаружены остатки каждого вида, от общего числа мест находок, и
относительное обилие - доля остатков каждого вида, выраженная в
процентах, по отношению к общему числу определенного материала.
Значительная часть материала представлена нижними челю-стями мелких
видов ночниц – Myotis daubentonii, М. brandtii,
M. ikonnikovii и М. frater.
|
пещеры
виды
|
Музейная
|
Разбойничья
|
Титюниха
|
Денисова
|
всего
|
n
|
n
|
п
|
n
|
|
Myotis
dasycmene
|
42
|
5
|
11
|
1
|
59
|
|
М. daubentonii
|
17
|
5
|
16
|
2
|
40
|
|
М. brandtii
|
11
|
4
|
4
|
n
|
19
|
|
М. frater
|
n
|
n
|
3
|
n
|
3
|
|
М. ikonnikovii
|
3
|
3
|
10
|
1
|
17
|
|
М. blythii
|
8
|
n
|
8
|
2
|
18
|
|
Plecotus
auritus
|
25
|
14
|
9
|
n
|
48
|
|
Eptesicus
nilssonii
|
15
|
11
|
3
|
2
|
31
|
|
Murina
leucogaster
|
n
|
n
|
1
|
1
|
2
|
|
Myotis
sp. small
|
105
|
14
|
16
|
6
|
141
|
|
всего
|
226
|
n
|
81
|
15
|
378
|
Таблица
3. Количественное соотношение остатков мелких ночниц в танатоценозах
голоценовых отложений пещер северо-западного Алтая.
|
виды
|
относительное обилие
|
встречаемость
|
n
|
%
|
n
|
%
|
|
Myotis dasycmene
|
59
|
15,6
|
4
|
13,8
|
|
М. daubentonii
|
116
|
30,7
|
4
|
13,8
|
|
М. brandtii
|
63
|
16,7
|
4
|
13,8
|
|
М. frater
|
4
|
1,1
|
1
|
3,4
|
|
М. ikonnikovii
|
37
|
9,8
|
4
|
13,8
|
|
М. biythii
|
18
|
4,8
|
3
|
10,3
|
|
Plecotus auritus
|
48
|
12,7
|
3
|
10,3
|
|
Eptesicus nilssonii
|
31
|
8,2
|
4
|
13,8
|
|
Murina leucogaster
|
2
|
0,5
|
2
|
6,9
|
|
всего
|
378
|
100,0
|
29
|
100,0
|
Таблица
4. Относительное обилие и встречаемость остатков разных видов рукокрылых
в голоценовых отложениях пещер северо-западного Алтая.
Mopфологическивидовуюпринадлежностькаж-дой нижней
челюсти нам определить не удалось. По этой причине процентное соотношение
краниальных остатков этих таксонов в ориктоценозах каждой из пещер
было рассчитано без учета нижних челюстей, на основании предварительно
определенных черепов и верхнечелюстных элементов (табл. 3). Затем,
в соответствии с долевыми показателями, было рассчитано конечное
число остатков каждого вида от общего числа остатков (в том числе
неопределимых нижних челюстей). И уже для полученных показателей
числа остатков каждого вида мелких ночниц были рассчитаны встречаемость
и относительное обилие.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1. Структура и состав голоценового сообщества
пещерных рукокрылых Алтая.
Субфоссильные кости рукокрылых имеют разную сохранность и окраску.
По-видимому, анализируемый материал накапливался в течение голоцена
довольно продолжительное время, вплоть до современности. Точно установить
возраст ископаемых остатков не представляется возможным.
Основная масса субфоссильных остатков имеет нативную окраску костей
– белую или пепельно-серую. Редко можно встретить кости, имеющие
рыжую окраску, иногда с черными прокрасами марганца. Бурый и рыжеватый
оттенок костей чаще всего обусловливается наличием в породе трехвалентного
железа.
В голоценовом танатоценозе нами установлено присутствие остатков
девяти видов рукокрылых: водяной ночницы Myotis daubentonii,
ночницы Брандта М. brandtii, прудовой ночницы М. dasycneme,
длиннохвостой ночницы М. frater, ночницы Иконникова М.
ikonnikovii, остроухой ночницы М. blythii, бурого ушана
Plecotus auritus, северного кожанка Eptesicus nilssonii
и сибирского трубконоса Murina leucogaster. Морфологически
все полуископаемые материалы полностью идентичны современным видам.
По показателю относительного обилия самым многочисленными в субфоссильном
сообществе рукокрылых являются мелкие виды ночниц: водяная и Брандта
(табл. 4). На втором месте находится прудовая ночница, на третьем
– бурый ушан, далее – северный кожанок, затем – остроухая
ночница. Малочисленным видом является ночница Иконникова, наиболее
редки в сборах длиннохвостая ночница и сибирский трубконос.
По встречаемости картина несколько иная (табл. 4). На первых местах
– все те же водяная ночница, ночница Брандта и прудовая ночница.
Однако к ним присоединяется сравнительно малочисленная ночница Иконникова.
По-видимому, это свидетельство того, что обычно этот вид не образует
больших скоплений. Костные остатки северного кожанка также обычны.
На втором месте находятся бурый ушан и остроухая ночница. Наши исследования
современной и полуископаемой фауны Chiroptera Горного Алтая показали,
что в голоценовое время остроухая ночница на исследуемой территории
была распространена гораздо шире, чем в современности. В ходе трехлетнего
исследования в тех пещерах, где были собраны субфоссильные кости
этой крупной ночницы, живые зверьки нами обнаружены не были. Единственная
находка современной крупной колонии остроухих ночниц сделана юго-западнее
от места наших исследований – в Чарышском районе на правом
берегу р. Чарыш, около пос. Усть-Чагирка (Розен 1953; Стрелков,
1968; Горетовская и др., 2002). Остатки сибирского трубконоса по
встречаемости находятся на предпоследнем месте. Длиннохвостая ночница
— самый редкий вид в голоценовом танатоценозе. Полученные показатели,
по-видимому, близки к истинным значениям распространенности и численности
данного вида на территории современного Алтая. Исследования современной
фауны региона показали, что М. frater проявляет высокую избирательность
по отношению к пещерам, единственная крупная колония этого вида
найдена только в Старокаракольской пещере. В пещере Титюниха, из
которой собраны голоценовые остатки этого вида, в настоящий момент
длиннохвостая ночница не найдена.
2. Изучение начальных этапов фоссилизации остатков рукокрылых
в условиях пещер.
Пещерные захоронения рукокрылых по приведенной выше классификации
относятся в первому типу. Как правило, концентрация костного материала
летучих мышей в пещерах происходит без участия каких-либо дополнительных
факторов, кроме гибели зверьков и их последующего захоронения. Чаще
всего переотложение материала не происходит, и мы имеем дело с редким
случаем первичного захоронения тушек погибших летучих мышей в месте
гибели. Однако в более влажные климатические периоды остатки рукокрылых
более древних, плейстоценовых захоронений могли быть перенесены
водными потоками в пределах полости пещеры и таким образом переотложены.
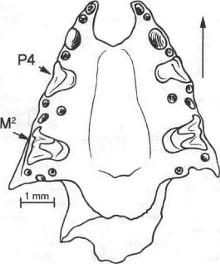 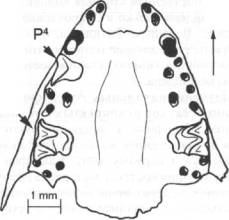
Рис.
2. Фрагментированный череп Plecotus auritus из пещеры Разбойничья.
Рис. 3. Фрагментированный череп Eptesicus nilssonii из пещеры
Разбойничья
В различные периоды жизненного цикла рукокрылых
характер расположения погибших животных будет отличаться. Так во
время зимовок значительная часть погибших зверьков будет концентрироваться
в одном месте пещеры, поскольку большинство видов в этот период
проявляет колониальность. Особи, погибшие в летний период, нередко
хаотично расположены в привходовой части пещеры. Влажностно-температурные
условия в полостях пещер существенно не изменяются на протяжении
сезона, хотя и подвержены некоторым флуктуациям в зависимости от
климата снаружи. Однако микроклиматические условия в разных частях
пещеры могут значительно отличаться, поэтому зверьки в полости пещеры
локализуются избирательно. Разные виды предпочитают различные типы
микроклимата и микрорельефа, проявляя значительную долю консерватизма,
и неравномерно распределяются в пещерном пространстве (Стрелков,
Ильин, 1990; Смирнов и др., 1999; Томиленко, 2002). Соответственно,
трупики погибших животных распределены также неравномерно. Остатки
рукокрылых, проявляющих тяготение к пещерным трещинам и щелям в
стенах (трубконосы), в значительно меньшей степени представлены
в поверхностном слое грунта, но концентрируются в субстрате трещин
и щелей. Напротив, тушки высококолониальных рукокрылых (например,
большинства ночниц), предпочитающих располагаться открыто массовыми
скоплениями, будут падать на пол пещеры и здесь же переходить в
ископаемое состояние. Таким образом, пространственное распределение
ископаемых остатков рукокрылых по горизонтали, прослеженное в вертикальной
колонке пещерных отложений, может играть важную роль в процессах
познания поведенческих особенностей видов летучих мышей, а также
объяснить таксономический набор и соотношения рукокрылых в ориктоценозах.
Во многом облик ориктоценоза определяется морфологическими особенностями
объектов захоронения. Например, основная масса плейстоценовых материалов
по летучим мышам представлена только разрозненными зубами, которые
сложно идентифицировать. Анализ субфоссиль-ных материалов показал,
что разные элементы зубного ряда с разной продолжительностью сохраняются
в челюстях при фоссилизации. Вероятно, это отражается в конечном
составе ориктоценозов, где разные члены зубного ряда имеют разное
долевое участие.
Соединительная ткань, которая удерживает зубы летучих мышей в альвеолах,
разрушается (или размягчается) чрезвычайно быстро, и, кроме корней,
зубы в альвеолах уже ничего не удерживает. Экспериментально установлено,
что зубы свободно вынимаются из челюстей после пятиминутного нахождения
в воде при температуре 80-90 °С. У тушек с разрушающимися мягкими
тканями, даже при незначительном перемещении, из челюстей сразу
же выпадают зубы. Однако, если череп находился в субстрате с цементирующими
свойствами, в определенной влажности, происходит закрепление зубов
в челюсти: субстрат заполняет промежутки между зубами и препятствует
их выпадению. В этом случае челюсть переходит в ископаемое состояние
с полным зубным рядом.
Для верхнего зубного ряда рукокрылых установлено, что не все зубы
выпадают из челюстей одновременно: первыми выпадают однокорневые
одновершинные зубы (клыки, премоляры и резцы), которые плохо закреплены
в альвеолах и имеют простой стержневидный корень. Двух-трехкорневые
зубы – премоляры и моляры, остаются в челюстях дольше. Из моляриформных
зубов первыми выпадают третьи моляры, особенно у видов с редуцированными
М3 (например, у ушана, М3 которого удерживается фактически только
двумя корнями, а третий корень располагается практически на одной
линии с двумя другими и плохо укрепляет коронку к челюсти). Чем
менее сжат в передне-заднем направлении М3, тем он более прочно
укреплен своими корнями в челюстной кости (например у ночниц). После
третьих моляров выпадают остальные. Последовательность их выпадения
также зависит от формы, расположения корней и положения в челюстной
кости. Как показывают наблюдения, М2 дольше остается в челюсти,
чем М1 (рис. 2, 3). По-видимому, это определяется положением этих
близких по форме зубов в вернечелюстной кости: М2 расположен у основания
скуловой дуги, и его корни, уходя в кость глубже, чем у М1, лучше
в ней закреплены. Помимо этого череп рукокрылых со стороны назальных
костей сильно подвержен разрушению. Соответственно, стержневые корни
М1 быстрее обнажаются, и зуб быстрее и легче выпадает. Дольше всех
в челюсти сохраняются верхние большие премоляры Р4 (рис. 2, 3),
так как они часто имеют сильно искривленные корни, удерживающие
зубы в альвеолах.
Особенности пещерных условий вызывает ряд специфических процессов,
сопровождающих фоссилизацию костного материала. Наблюдения показали,
что в лужицах пещерной воды субфоссильные кости летучих мышей быстро
становятся очень хрупкими и способны полностью раствориться. Первыми
растворяются ажурные части черепной коробки и осевого скелета. Более
устойчивыми являются трубчатые кости, ростральные части черепа и
нижние челюсти. Не исключено, что в конечном итоге остаются только
зубы, как части скелета наиболее устойчивые к подобной эрозии.
Основная масса субфоссильного материала по рукокрылым скапливается
в небольших трещинах в стенах пещер, а также на полу, под излюбленными
местами в микрорельефе пещеры, в которых происходит концентрации
животных во время зимовок или в летний период. В табл. 1 обозначен
весь спектр местонахождений, в которых были собраны кости млекопитающих.
В пробах РАЗ-1 и МУЗ-2 (табл. 1 и табл. 5) из пещер Разбойничья
и Музейная материал, по-видимому, частично переотложенный водными
потоками, представлен целыми и сильно поврежденными черепами, нижними
челюстями и частями осевого скелета. Образцы МУЗ-4-8, ТИТ-1 и ТИТ-3
собраны в трещинах в стенах пещер. Здесь материал довольно сильно
поврежден. Пробы ДЕН-1, МУЗ-1 и МУЗ-3 собраны в поверхностном слое
грунта. Две последние пробы содержат сильно фрагментированные остатки
рукокрылых. Однако в их разрушении немалую роль сыграл человеческий
фактор, так как значительная часть костного материала подвергается
механическому разрушению во время посещений пещер туристами и спелеологами.
|
пробы
|
всего остатков
|
целые черепа
|
поврежденные черепа
|
|
п
|
%
|
п
|
%
|
| РАЗ-1 |
57
|
8
|
14,0
|
12
|
21,1
|
| ТИТ-1 |
5
|
1
|
20,0
|
—
|
—
|
| ТИТ2 |
19
|
4
|
21,1
|
4
|
21,1
|
| ТИТ-3 |
4
|
—
|
—
|
—
|
—
|
| ТИТ-4 |
21
|
—
|
—
|
1
|
4,8
|
| МУЗ-1 |
45
|
1
|
2,2
|
7
|
15,6
|
| МУЗ-2 |
4
|
1
|
25,0
|
1
|
25,0
|
| МУЗ-3 |
76
|
—
|
—
|
3
|
3,9
|
| МУЗ-4 |
41
|
—
|
—
|
—
|
—
|
| МУЗ-5 |
23
|
1
|
4,3
|
4
|
17,4
|
| МУЗ-6 |
23
|
3
|
13,0
|
1
|
4,3
|
| МУЗ-7 |
25
|
2
|
8,0
|
—
|
—
|
| МУЗ-8 |
8
|
—
|
—
|
1
|
12,5
|
| МУЗ-9 |
13
|
—
|
—
|
1
|
7,7
|
| ДЕН-1 |
6
|
2
|
33,3
|
—
|
N
|
| ДЕН-2 |
8
|
—
|
—
|
—
|
—
|
|
всего
|
378
|
23
|
6,1
|
35
|
9,3
|
Таблица
5. Количественное соотношение целых и поврежденных черепов рукокрылых
в голоценовом ориктоценозе, представленном краниальными остатками.
Галереи Денисовой пещеры не представляют особого
интереса для туристов, посещаются ими значительно реже и поэтому
материал здесь сохраняется лучше (ДЕН-1). Сильно поврежден материал
в пробе МУЗ-9. Субстрат этой пробы представлен перемешанным грубообломочным
материалом кальцитовых кристаллов, среди которых обнаружены сильно
поврежденные остатки краниального и осевого скелета рукокрылых.
Таким образом, сохранность костного материала рукокрылых зависит
от состава, структуры и интенсивности перемещивания отложений вмещающей
породы.
БЛАГОДАРНОСТИ
Автор сердечно благодарит А.К. Агаджаняна за всестороннюю
поддержку проведенного исследования, и А. Макивского - за
существенную помощь в сборах суб-рецентных материалов в пещерах
Алтая. Искренняя признательность СВ. Крускопу и А.В. Лопатину за
ценные замечания по содержанию рукописи. Рис. 1 выполнен А.В. Абдульмановой.
Возможность проведения полевых работ на базе археологического лагеря
"Денисова пещера" Института Археологии и этнографии СО РАН предоставлена
М.В. Шуньковым.
Работа выполнена при поддержке ФЦП "Коэволюция экосистем в условиях
глобальных изменений прошлого", РФФИ (грант № 02-04-48458) и программы
"Историческая динамика биоресурсов как предпосылка оценки их современной
охраны и эксплуатации".
Литература
Агаджанян А.К. Комплексные биостратиграфические исследования: учебное
пособие / под ред. П. А. Каплина. М.: Из-во Моск. ун-та, 1987.107
с.
Горетовская О.С., Рыжков Д.В., Бурмистров М.В. Изучение фауны рукокрылых
на территории Алтайского края // Plecotus et al., pars spec. 2002.
С. 106-108.
Николаев СВ. Литология, геохимия, биостратиграфия, палеогеография
голоцена Денисовой пещеры // Денисова пещера. Ч. 1. Новосибирск:
ВО Наука, 1994. С. 207-245.
Оводов Н.Д. Пещерные зимовки летучих мышей в южных районах Сибири
// Зоол. проблемы Сибири. Новосибирск: Наука, 1972. С. 439-440.
Оводов Н.Д. Субфоссильные остатки рукокрылых в пещерах Сибири и
Дальнего Востока // Матер. I Всес. совещ. по рукокрылым. Л.: Изд-во
ЗИН АН СССР, 1974. С. 84-90.
Оводов Н.Д. Пещерная гробница алтайских пищух (Ochotona alpina
Pall.) // VI съезд Териол. об-ва. Тез. докл. М.: Изд-во РАН, 1999.
С. 180.
Розен М.Ф. Массовое скопление летучих мышей в одной из пещер Алтая
// Природа, 1953. № 9. С. 118-119.
Смирнов Д.Г., Курмаева Н.М., Вехник В.П. Динамика численности и
пространственное распределение зимующих рукокрылых (Chiropetra,
Vespertilionidae) в одной из штолен Самарской Луки // Plecotus et
al., 1999. № 2. С. 67-78.
Стрелков П.П. Остроухие ночницы на Алтае // Природа, 1968. № 2.
С. 59-61.
Стрелков П.П., Ильин В.Ю. Рукокрылые (Chiroptera, Vespertilionidae)
юга среднего и нижнего Поволжья // Тр. Зоол. ин-та АН СССР, 1990.
Вып. 225. С. 42-167.
Тиунов М.П. Рукокрылые Дальнего Востока России. Владивосток: Дальнаука,
1997. 134 с.
Томиленко А.А. Зимовка рукокрылых (Vespertilionidae) в Новосибирской
области // Plecotus et al., pars spec. 2002. С. 99-106.
Филлипов А.Г., Тиунов М.П. Остатки рукокрылых в пещерах Иркутской
области // Plecotus et al. 1999. № 2. С. 100-108.
|
